

Plan
Bergson opposait le temps à l’espace comme l’esprit à la matière, ce qui fait qu’il concevait la transition entre la rétention des événements dans le souvenir et l’orientation de l’habitude vers l’action comme une distraction de « notre attention à la vie ». Husserl fondait le sens d’être des événements et autres choses qui durent sur la pluridimensionnalité du processus de leur constitution subjective par l’activité rétentio-protentionnelle de la conscience intime du temps. Autant la conception traditionnelle d’un temps linéaire et d’un espace euclidien éloigne le premier, autant l’intuition du dynamisme pluridimensionnel de la temporalité rapproche le second des enseignements qu’on peut retirer de la découverte des fondements de la mémoire et de l’anticipation dans les cartes cognitives de la formation hippocampique émergeant des patterns d’activité des cellules de lieu, ou cellules de temps.
Un retour à Bergson ?
Quand les neurosciences veulent réhabiliter le corps contre l’intellectualisation de la cognition par la théorie des représentations mentales, elles revisitent plus volontiers Bergson que Husserl. Dans Matière et mémoire elles trouvent une théorie sensori-motrice de la perception où elles croient voir l’ébauche d’une biologie de l’incarnation qui reconnaît l’importance du système moteur (A. Berthoz 2008). Cette relecture neuronale semble aller à rebours des intentions de Bergson, qui voulait limiter le corps au présent des besoins de l’organisme afin de doter l’esprit d’un pouvoir de « survivance en soi du passé » dont la matière est dépourvue. Abandonnant aux psychologues la mémoire habitude et ses automatismes, il croyait épargner au « souvenir pur » toute contamination par l’ordre spatial des choses, du corps et du cerveau. Dès l’Essai sur les données immédiates de la conscience Bergson subordonnait l’espace comme extérieur et matériel à la durée assimilée à l’esprit. L’incorporation de l’esprit répondrait seulement aux exigences de l’action – non à des motifs désintéressés tels que la connaissance, la contemplation ou le rêve. Condescendant à se rendre utile dans la vie normale – une disposition qui cesse avec la pathologie – la mémoire puise en soi une sélection de souvenirs pertinents pour les tâches actuelles. Mais la fixation de l’événement personnel dans la singularité de son occurrence ne tiendrait qu’à notre être intérieur sans rien devoir à l’extérieur.
L’analogie champ temporel – champ visuel
Loin d’opposer le temps à l’espace, Husserl guide l’analyse de la conscience du temps sur l’analogie entre le champ visuel de la perception et le « champ temporel » de la conscience du temps. Pour la conscience perceptive, un objet dans l’espace se donne par esquisses ou aspects, dont un seul est clair et distinct, tandis que les autres aspects s’estompent en perspective (Ding und Raum, 1907). Pour la conscience du temps un objet temporel se donne « maintenant » dans un point de durée actuel, tandis que les points de durée écoulée reculent dans un passé toujours plus lointain (Zeitbewusstsein, 1905). Dans les deux cas la variété des modes d’apparition n’empêche pas mais fait qu’ils soient les modes d’apparition d’un objet identique. Même perspectivisme de l’être subjectif, qui est toujours situé et qui donne sens à ses objets en les rapportant au « ici et maintenant ».
La phénoménologie de la conscience du temps procède uniquement par réflexion sur les vécus, sous condition de réduction méthodique des présuppositions usuelles d’existence relatives à un temps du monde englobant les états de conscience. Ce n’est pas un modèle théorique en attente de validation empirique, mais un effort personnel de la part du phénoménologue pour remonter de l’évidence de son expérience des objets temporels aux conditions nécessaires a priori de leur constitution subjective. La phénoménologie du temps est donc la description de la structure transcendantale de la conscience d’un sujet percevant pour lequel il y a quelque chose de tel que des objets temporels dans le temps objectif (des choses durables, des événements, des processus ou des mouvements). Les objets qui durent acquièrent la place qui leur revient dans l’ordre chronologique dans la mesure où ils se manifestent à la conscience d’un sujet qui les saisit. D’où la contrainte de devoir présenter leurs phases successives dans le présent actuel de la perception, quitte à ce qu’elles disparaissent progressivement dans le flux héraclitéen des vécus de la conscience.
La conscience-temps est un champ de manifestation phénoménale pré-empirique, proto-temporel, autrement dit un a priori transcendantal. La configuration du champ spécifie les conditions nécessaires a priori pour l’apparition à un sujet percevant et pour la saisie active par ce sujet d’un objet temporel comme entité unique et identique à travers la variété de ses modes de présentation. Les places des objets dans le temps objectif et les intervalles entre les objets à leurs places respectives sont déterminés sur la base de l’expérience vécue de l’éloignement progressif des phases de la présentation perceptive de ces objets par rapport au maintenant actuel. Le temps objectif est un ordre fixe de places elles-mêmes fixes à des distances respectives absolument déterminées. Objectivement, « le vol d’un oiseau », considéré indépendamment du point de vue du sujet percevant, est un continuum de places fixes remplies d’un contenu qualitatif (comme dans l’Univers-bloc d’Einstein où passé, présent et futur sont également réels) :
« Objectivement, des relations fonctionnelles mathématiques fixes nommées « causalités mathématiques » existent dans un temps, également fixe. L’oiseau vole : une extension déterminée du temps objectif est objectivement remplie de telle et telle façon, et elle pourrait idéalement être remplie au sens du non-changement, alors qu’elle l’est au sens d’un changement : un vol. Le changement exprime un mode déterminé de remplissement d’un temps, le non-changement en exprime un autre. Mais dans le temps objectif, cette extension remplie est fixe » (Bernauer Manuskripte über Zeibewusstsein, N10, 182, 1917)1.
Un objet temporel est le corrélat d’un acte ou d’une suite d’actes de saisie active par le sujet percevant d’un matériau sensoriel comme constituant une entité unique et identique à travers ses modes de présentation variables pendant le cours complet de sa présentation. Exemples : on entend la mélodie et pas seulement une suite de sons. On entend le son d’une cloche, la note « do » jouée au violon, le sifflet de la locomotive. On observe le vol d’un oiseau. On voit passer un escadron de cavalerie au galop ; ou encore – pour être pour être plus explicite sur le contexte historique :
« Je me souviens des soldats qui défilaient hier en chantant la mélodie „Ich hatt‘ einen Kameraden“, et en même temps me viennent plusieurs souvenirs de soldats qui défilent en chantant ce même chant » (Bernau, Novembre 1917)2.
Le problème : une succession d’impressions n’est pas une impression de succession
Les « impressions » au sens n°1 sont des données de fait qui ne renvoient qu’à elles-mêmes, des data, qui ou bien sont dans l’actualité effective, ou bien ne sont plus. « L’impression » au sens n°2 est déjà une conscience de quelque chose, un acte intentionnel orienté vers un objet dans une tension qui déborde l’actuel vers un possible. Comment est-il possible de passer de la succession d’impressions à l’impression de succession ? La morphologie dynamique de la conscience-temps rend possible la transition. L’activité de la conscience-temps change la succession en objet temporel, un processus qu’on peut décomposer comme suit :
– La saisie du maintenant qualitativement rempli et sa constante réactualisation au foyer-source de la perception ;
– La modification comme « juste passé » du maintenant repoussé hors du foyer de la perception effective actuelle ;
– La rétention du maintenant modifié comme horizon de juste passé du maintenant actuel de la présentation du même objet ;
– L’enveloppement de la structure : foyer perceptif-horizon rétentionnel du maintenant actuel dans un nouvel horizon du « juste passé du juste passé », et ainsi de suite ;
– L’acte de visée d’identité qui traverse tous les horizons rétentionnels et sauvegarde l’unité de l’objet temporel...
Le continuum des horizons rétentionnels du maintenant actuel et de chaque phase antérieure d’activité de conscience conserve – par le fait même que ces horizons sont régulièrement emboîtés les uns dans les autres – l’héritage de la présentation initiale de l’objet temporel. Sa structure eidétique : actes de détermination du maitenant + transition continue du maintenant actuel au maintenant juste passé, et ainsi de suite, jusqu’au maintenant initial. Cette structure est celle de "variété continue" (stetige Mannigfaltigkeit de Riemann 1854) : concept général de grandeur à n dimensions simplement définie par la possibilité de la transition continue d’un mode de détermination à un autre. La régularité de cet ordre d’emboîtement des horizons sauvegarde la prise de la conscience sur l’objet, dans la mesure où la transition continue d’un horizon à l’autre maintient ouvert le chemin d’une visée intentionnelle toujours possible, qui prendrait cet objet comme cible.
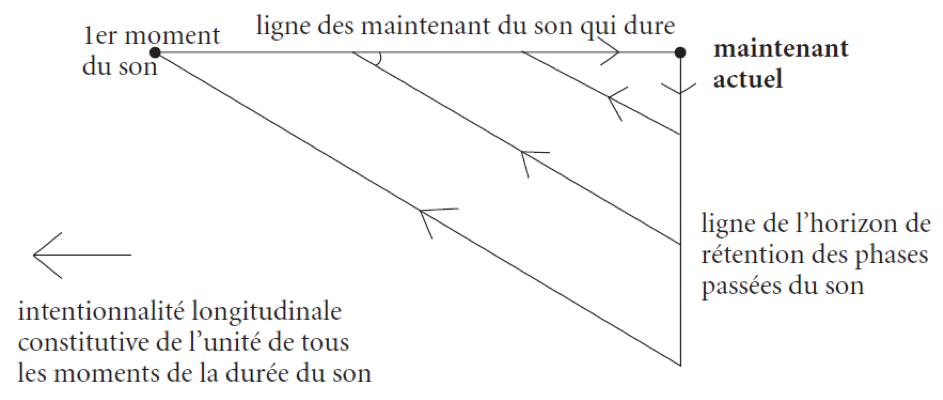
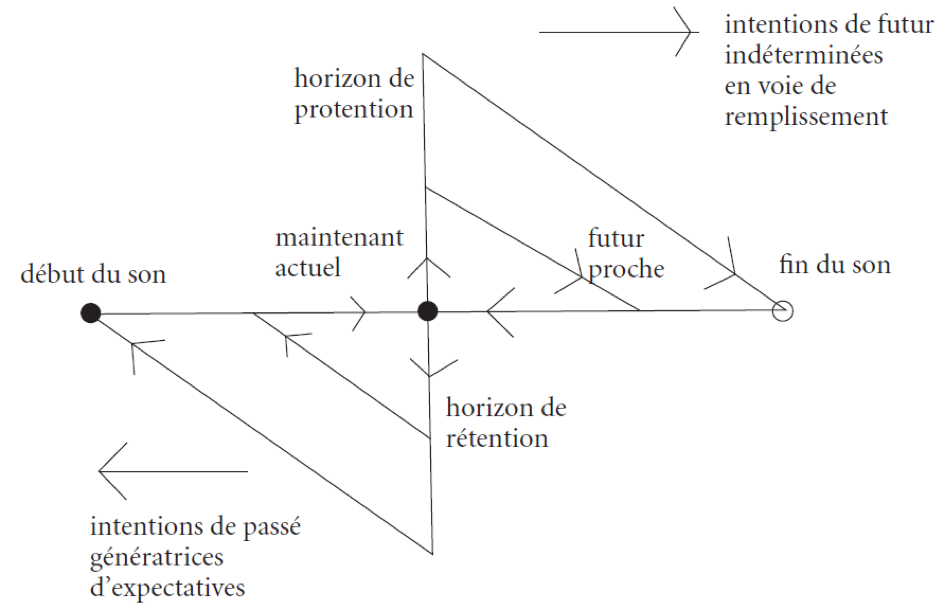
Justification de la protention
L’impression originaire n’a de signification intentionnelle que si elle répond à une attente relativement déterminée, où le sujet percevant va au-devant de ce qui arrive avec une prédisposition à l’accueillir « les bras ouvert ». Le champ proto-temporel de la conscience-temps se révèle d’une structure complexe pluridimensionnelle : le continuum des horizons rétentionnels doit être replongé dans le champ des expectatives du sujet concernant l’événement futur. Ce champ est structuré avec un foyer-source de l’événement survenant et une série d’horizons d’expectatives, enveloppant ce foyer et enveloppés les uns dans les autres, qui va du plus large et moins déterminé au plus étroit et plus précis. La protention est l’intention, symétrique de la rétention, avec laquelle le sujet percevant cherche à satisfaire les exigences de saturation des lacunes qui subsistent dans la détermination de ses expectatives, en se projetant toujours à nouveau vers l’événement prochain comme potentiellement satisfaisant.
Un horizon d’expectative absolument vide ou indéterminé n’est l’attente de rien ; complètement rempli, il n’est plus attente mais intuition de l’objet perçu. Comment l’horizon protentionnel peut-il être déterminé si ce n’est par une première présentation de l’objet : d’où un objet inconnu ne sera jamais perçu ? La solution : s’en tenir à la constitution de la temporalité de l’objet actuellement donné par auto-application continue (recouvrement) d’un continuum de petites différences inchoatives et juxtaliminaires (différentielles de temps). Dans un dévoilement progressif de la structure complète de la conscience-temps, la possibilité de la détermination du champ protentionnel se révèle dépendre d’une projection de la stratification d’horizon des rétentions sur la dimension de « l’à venir », comme retour de « la même chose » selon le même mode. Le champ proto-temporel est rétentio-protentionnel.
Pour un rapprochement de la phénoménologie de la conscience du temps et des neurosciences il faut se garder de subordonner le proto-temps constituant de la phénoménologie au temps constitué chronométrique des processus oscillatoires cérébraux. Mais comment est-ce possible, compte tenu du fait que, comme toute science de la nature, les neurosciences sont des sciences de la mesure : mesure des oscillations cérébrales en électroencéphalographie, mesure du flux sanguin cérébral en imagerie cérébrale ? Une part de la recherche des bases neurales de notre expérience du temps s’expose à la critique phénoménologique parce qu’elle se contente de présupposer un temps constitué sans chercher à remonter aux origines subjectives du temps. Héritage de la tradition psychophysique, le recours à des tâches de comparaison et d’estimation des intervalles de temps ne donne accès qu’à un temps métrique qui ne sollicite le sujet que comme porteur d’une horloge interne, centrale ou localement distribuée. En dernière analyse, on rend compte des propriétés métriques des intervalles de temps perçus avec les propriétés métriques de l’activité des cellules nerveuses : potentiels d’action, fréquence, amplitude, pente, durée d’oscillation. À part la différence d’échelle, un temps chronométrique renvoie toujours à du temps chronométrique. Mais, indépendamment de toute idée de mesurer des intervalles de temps, le phénomène même de la possibilité du temps, qui dépend de la mise en ordre de succession des moments, ou phases successives d’un événement ou d’une chose durable n’est pas encore mis au jour. Or, c’est précisément sur la géométrie des propriétés formelles a priori des processus temporels (la relation d’ordre de succession linéaire de leurs phases successives) que portent les diagrammes de la constitution du temps dans la conscience. Pour une neuroscience du temps qui dépasserait la simple mesure des intervalles de temps ces diagrammes pourraient être le bon niveau stratégique d’un dialogue avec la phénoménologie.
La phénoménologie remonte aux origines subjectives du sens du temps en une expérience de pensée du type cogito, où chacun est invité à réactiver les actes de conscience qui constituent les objets temporels. Les neurosciences cognitives considèrent le sens du temps comme une propriété du comportement qui doit avoir des corrélats localisés dans des structures du cerveau et s’expliquer par mécanismes fonctionnels. La première approche est immanente : elle sollicite nos activités et capacités, notre expérience vécue et la réflexion sur ce vécu. L’autre approche est transcendante : elle aborde l’objet « temps » par la voie indirecte du modèle théorique (carte cognitive de Tolman 1948) et de l’expérimentation sur modèle animal (rat dans un labyrinthe) grâce aux progrès de la technologie d’enregistrement intracellulaire (tetrode recording).3
L’hippocampe : carte cognitive interne
La courte histoire des neurones de temps s’inscrit dans le prolongement de l’histoire des neurones de lieu qui commence avec le neurophysiologiste J. O’Keefe, prix Nobel 2014 pour la découverte dans l’hippocampe du cerveau des mammifères de cellules à champ récepteur spatial, bases neuronales du sens de la position de l’individu dans l’espace environnant. Interprétée par la théorie des cartes cognitives spatiales de Tolman et Hebb (1949), cette découverte contredit la thèse d’un cerveau sensori-moteur et non organe de représentation. La fonction de l’hippocampe comme base de la mémoire et de l’individuation des souvenirs personnels par un processus spatialisant suggère, contre Bergson, que l’actualité de l’état cérébral ne fait pas obstacle à la conservation du passé. Le rôle de l’hippocampe dans le traitement de l’information cognitive serait de réemployer la carte cognitive spatiale pour l’organisation de l’expérience individuelle selon des dimensions autres que spatiales : plus particulièrement, l’organisation des moments d’un épisode selon leur ordre de succession dans le temps.
Comme illustration, on se rapportera à une étude pionnière par O’Keefe et Dostrovsky, (Brain Res.1971). Le rat était simplement maintenu avec la main dans une position et une orientation déterminées. L’enregistrement était encore unicellulaire. L’histogramme des fréquences de décharge d’une cellule unique (dans la région CA1) pendant chaque position et orientation révélait les préférences d’activation du neurone : la cellule décharge toutes les fois que le rat se trouve dans une certaine position. Dans cette étude pionnière la découverte d’une activité neuronale uniquement liée à la localisation de l’animal se dégage encore mal des influences de l’orientation et de la stimulation tactile.
Le maintien passif dans une position donnée ne permettant pas de déterminer l’usage de cette indication de position par rapport au comportement de l’animal en contexte écologique, les travaux ultérieurs allaient introduire une tâche imposée pendant l’enregistrement. C’est ainsi qu’en une étude ultérieure O’Keefe et Speakman (Exp. Brain Res.1987) ont pu démontrer la relativité du champ de lieu des cellules de lieu par rapport aux indices du but de l’animal. La tâche était de choisir la branche d’un labyrinthe en croix contenant la récompense en présence de certains indices visuels (tâche de perception), ou en leur absence (tâche de mémoire). Une majorité de cellules ont un champ de lieu solidaire des indices mobiles du but. Pour une minorité de cellules le champ de lieu reste invariable par rapport à des indices statiques du fond. On note la similarité, en particulier la fréquente superposition, des champs de lieu de chaque cellule dans la tâche de perception et dans la tâche de mémoire. Interprétant ces observations en termes de carte cognitive, les auteurs avancent que l’orientation préférée par rapport au fond procure à l’animal une carte par défaut qu’il réoriente à l’aide des indices mobiles du but manipulés par l’expérimentateur.
La mise en évidence de l’importance de la forme géométrique de l’environnement pour la localisation du champ de lieu des cellules de lieu de l’hippocampe est le fait d’un travail encore ultérieur où O’Keefe et Burgess (Nature 1996) avaient placé le rat dans une boîte au format modifiable. Les cartes des taux d’activité des cellules de lieu enregistrées dans les régions CA1 et CA3 montrent clairement que lorsque la boîte passe d’un format à un autre : grand carré de 122 x 122 cm, petit carré de 61 x 61 cm, et de là à un format rectangulaire de 61 x 122 cm, l’emplacement du pic d’activité par rapport aux murs reste pratiquement inchangé. Sur la base de cette constatation (limitée à la géométrie du carré et du rectangle), les auteurs croient pouvoir inférer une règle générale de la formation du champ de lieu. Le pic d’activité de chaque cellule de lieu étant localisé à une distance fixe des murs, la place et la forme du champ dépendent donc de la forme de la boîte : mathématiquement, le champ résulterait d’une somme de fonctions gaussiennes de la distance du rat par rapport à chacun des murs de la boîte. Cette généralisation était, toutefois, prématurée dans la mesure où les changements de format de boîte expérimentés n’étaient pas ceux qui peuvent entraîner une redéfinition de la carte cognitive, une éventualité que la poursuite des recherches devait confirmer.
Dans une étude plus récente, Wills et al. (Science 2005) ont systématisé la modification de la forme géométrique de l’environnement pour mettre à l’épreuve la prétendue fixité de sa représentation dans les cartes cognitives. L’exposition à une enceinte alternativement carrée et circulaire – cette fois – induit, au moins pour un groupe non négligeable de cellules de lieu, une recartographie rapide des représentations de l’environnement. Loin que les cellules enregistrées réagissent d’une manière uniforme, pour certaines la carte est insensible au changement, tandis que d’autres font la différence entre les deux configurations, d’emblée ou progressivement et que d’autres encore convergent vers une même représentation. Selon les auteurs, cette plasticité des cartes cognitives de l’hippocampe, rendant possible la transition d’un format de carte à un autre, pourrait expliquer l’aptitude de la mémoire à différencier les souvenirs d’événements similaires survenus dans des contextes différents. L’hippocampe, au filtre du concept spatial de carte cognitive, fait figure de substrat de la mémoire des événements en contexte (mémoire épisodique).
Les neurones de grille : système de triangulation topographique
Aux neurophysiologistes M.-B. Moser et E. Moser, colauréats avec O’Keefe du prix Nobel 2014, on doit la découverte dans le cortex entorhinal, structure d’entrée de l’hippocampe, des cellules d’un système de coordonnées géométriques interne pour la détermination des positions de l’individu dans l’espace. En matière de sophistication de la représentation de l’espace il y a là plus que le marquage sur une carte interne d’un lieu particulier traversé par l’animal dans sa course. Le pattern d’activation des cellules individuelles est organisé de façon à projeter sur l’environnement un grillage de triangles équilatéraux dont les sommets coïncident avec les maxima des taux de décharge de chaque cellule : comme si la cellule nerveuse déployait une carte topographique sur laquelle on peut localiser en termes de position, distance et orientation n’importe quel détail de l’environnement pourvu qu’on dispose d’un point de repère fixe pour le calibrage de la carte !
Les cellules de grille du cortex entorhinal sont décrites dans un article par Hafting et al, (Nature 2005) qui présente les données d’enregistrement de deux cellules du cortex entorhinal, l’une de la partie dorsale, l’autre de la partie ventrale. Pour chaque cellule la trajectoire de la course du rat dans une boîte carrée de 1 m. de côté est représentée par un tracé continu très enchevêtré avec les pics d’activation de la cellule superposés en une curieuse distribution triangulaire : la cellule décharge chaque fois que l’animal passe par le sommet d’un des triangles de base d’une grille virtuelle couvrant la surface de l’environnement. En regard, on donne les cartes des taux d’activation révélant un pavage de l’espace entier par de nombreux champs de décharge discrets localisés aux sommets de triangles équilatéraux juxtaposés en hexagone régulier. Les autocorrélogrammes de ces cartes démontrent la grande régularité de ce pavage de l’espace. On constate également une différence de finesse du maillage d’une région à l’autre, qui suggère que la représentation de l’espace par les grilles de la région dorsale du cortex entorhinal possède une résolution supérieure à celle des grilles de la région ventrale.
On a vu que les cellules de lieu de l’hippocampe du rat peuvent réorganiser entièrement la localisation de leur champ (recartographie) lorsqu’on change de forme la boîte. Globale au niveau de l’hippocampe, cette réorganisation coïncide avec un simple déplacement ou un simple pivotement du champ de grille des cellules du cortex entorhinal, qui sauvegarde la régularité périodique de sa structure triangulaire répétitive (Fyhn et al, Nature 2007). Quand la forme géométrique de la boîte n’est pas modifiée, mais que l’on change simplement la couleur des parois, il en résulte une modification du taux de décharge des cellules de lieu de l’hippocampe. En revanche, les grilles des cellules du cortex entorhinal restent invariables. De la comparaison du mode d’activité des cellules des deux régions les chercheurs infèrent la règle de fonctionnement du système cellules de lieu – cellules de grille : le codage neural du lieu se réalise de façon complémentaire (1) par actualisation d’un cadre universel rigide représenté par le champ de grille des cellules de grille (2) par attribution d’un code spécifique pour chaque circonstance par le champ de lieu des cellules de lieu.
Cellules de lieu ou cellules de temps ?
C’est dans l’optique de la recherche des bases neurales de la mémoire de la trajectoire suivie par le rat dans une tâche de navigation spatiale qu’a été mise en évidence l’existence de cellules sélectivement activées par un ou plusieurs des lieux traversés par l’animal dans sa course. Cette sélectivité d’activation n’est pas acquise d’emblée, mais repose sur une familiarité suffisante de l’animal avec l’environnement une fois la tâche apprise. Dans cette mesure, le phénomène “neurone de lieu”, couramment interprété en termes de carte cognitive spatiale, ne ressortit pas moins de la dimension temporelle, quoique de manière implicite. La question pour Pastalkova et al. (Science 321, 2008) était de la rendre explicite, ce à quoi une simple manipulation de la tâche expérimentale s’est avérée suffisante : le rat court sur place dans une roue en attendant de poursuivre sa course alternativement à droite et à gauche dans le labyrinthe dans un ordre de succession imposé par la tâche. Ce qu’on a découvert, c’est que de nombreuses cellules de l’hippocampe étaient actives dans les deux conditions, quoique dans un ordre séquentiel différent : les mêmes cellules de lieu dont le champ de décharge était ancré sur un certain emplacement dans le labyrinthe avaient leur « champ d’épisode » ancré chacun sur une même phase temporelle particulière de la course sur place dans la roue, et ce de façon récurrente à tous les essais. Illustration de la capacité du cerveau de générer une activité séquentielle, aussi bien dans la dimension du temps que dans celle de l’espace, les pics de décharge des 30 neurones simultanément enregistrés dans l’hippocampe s’ordonnent spontanément de manière à couvrir la totalité des phases temporelles de la course dans la roue. Chaque moment de cette durée étant caractérisé par l’activité d’une assemblée de neurones particulière, il est tentant d’interpréter la situation par analogie avec les neurones de lieu et le rappel des lieux traversés dans le labyrinthe, comme le rappel de l’ordre de succession des épisodes de la course sur place, entre le moment initial de l’entrée dans la roue et le moment terminal de l’arrêt de la roue et de l’accès au poste d’eau en guise de récompense. D’où, l’expression “champ d’épisode” et “cellules d’épisode”, qui suggère que cette capacité de mise en ordre séquentiel de l’activité d’une population neuronale fonde ici l’expérience du temps de la même manière qu’elle fonde là celle de l’espace.
L’usage du concept spatial de champ évoque l’existence d’un stimulus externe (ou d’un signal intéroceptif), tel que son entrée dans le champ récepteur d’une cellule dont c’est le stimulus préférentiel suffit à déclencher la décharge de cette cellule. Or, si l’environnement spatial (ou le milieu interne) peut éventuellement procurer le contenu sensoriel requis comme condition d’activation d’une cellule, il n’y a rien de tel dans le contexte temporel. Car, ou bien l’événement a lieu dans le moment présent de la perception, ou bien il est passé et il n’y a aucun contenu sensoriel qui puisse entrer dans le champ récepteur d’un neurone. C’est ainsi que l’on retrouve sur le plan neuronal le dilemme (sinon la solution au dilemme) de Brentano-Husserl de la conscience du temps. Là où le phénoménologue en appelle à l’horizon intentionnel de la rétention du maintenant juste passé pour soutenir la visée d’identité permanente de l’objet temporel à travers ses phases d’écoulement, le chercheur empirique manipulera la tâche comportementale de façon à en soustraire les paramètres de localisation, mouvement, distance parcourue, etc. et à ne retenir que l’ordre séquentiel des phases du comportement. Les variables spatiales à soustraire sont celles qui motivent la description de l’activité neuronale en termes de champ, parce qu’elles dénotent l’influence d’indices sur cette activité : indices de l’environnement pour la localisation, flux optique pour le mouvement, indices proprioceptifs pour la distance parcourue. En revanche, c’est uniquement par dérivation à partir de ces mêmes variables spatiales qu’on parlera de « champ temporel » pour les conditions d’activation des cellules de temps de l’hippocampe. Pour expliquer la récurrence d’un essai à l’autre de la préférence d’activation d’une cellule pour une même phase de la course, faute d’indices des phases de la durée, on ne peut pas l’attribuer à l’influence d’un élément concret de l’environnement, on ne peut que l’imputer “à la dynamique intrinsèque du réseau”.
Exemple de cette approche des neurones de temps par soustraction des dimensions spatiales dans une activité où se mélangent les influences de toutes les variables pertinentes pour la tâche, Kraus et al. (Neuron 78, 2013) reprennent le protocole de la course sur place avec une modification. Pendant le délai entre deux parcours du labyrinthe en sens alterné le rat court sur place sur un tapis roulant à une vitesse dont la variation aléatoire permet de dissocier la distance parcourue et le temps passé sur le tapis. Comme déjà vu, une majorité de neurones de l’hippocampe s’activent à une phase déterminée de la course sur place. Et les champs de décharge temporels contigus de l’ensemble des neurones couvrent la totalité de la période de délai. Quoique la majeure partie des cellules enregistrées soient influencées par les deux variables, certains neurones répondent plutôt au temps écoulé qu’à la distance parcourue. De ces “cellules de temps” les auteurs concluent qu’elles représentent, ou encodent le temps que le rat a passé sur le tapis roulant, tout en concédant la relativité de cette représentation du temps au contexte très particulier de la tâche, puisque les mêmes cellules ont un champ de lieu classique dans le labyrinthe. Les cellules à champ d’activation focalisé de l’hippocampe ne sont d’ailleurs pas les seules à manifester cet opportunisme de la représentation occasionnelle de la dimension temporelle en fonction du contexte de la tâche. Répliquant le protocole de course sur place sur tapis roulant en enregistrant, cette fois, les cellules de grille du cortex entorhinal, la même équipe (Kraus et al. Neuron 2015) a montré qu’avec leur pattern d’activation fort différent de celui des neurones de l’hippocampe, cette famille de cellules pouvait également représenter le temps. En effet, des cellules de grille déchargent à un moment particulier de la course sur place, et leurs champs de décharge sont distribués sur la totalité de la durée de la course. On retrouve dans la course sur place les mêmes patterns de décharge à pics multiples et organisation périodique, excepté qu’ils ne sont pas superposés aux patterns en grille caractéristiques de ces cellules dans la course en open field. Ce qui suggère un mécanisme de codage unique pour la trajectoire spatiale et pour la durée de course sur place. Le système des cellules de lieu et des cellules de grille de la formation hippocampique assurerait donc de la même manière la représentation cartographique de l’espace parcouru par l’animal et l’organisation chronologique de la mémoire épisodique de son expérience.
Qu’est-ce qui nous rend capables de saisir le maintenant ?
La sélectivité d’activation des neurones de l’hippocampe pour chacun des moments d’un intervalle de temps entre début et fin d’un épisode est peut-être la condition de possibilité matérielle de l’effectivité de la saisie du maintenant. Sans le profil de décharge unique d’une cellule de temps codant une phase particulière de la durée de l’objet temporel un acte de conscience pourrait-il déterminer un point de temps dans la suite continue des moments de cet objet ? L’attention au présent serait-elle assez sélective pour isoler en un point le contenu hylétique d’un acte de saisie si la conscience constituante du temps ne dérivait pas cette structure de point d’une géométrie intrinsèque du vivant ? Dans l’information cognitive sur le contexte spatiotemporel qui s’échange entre les aires pariétale et frontale l’hippocampe segmente des unités discrètes et contigües entrant dans le champ temporel des cellules de temps : le secret de la transformation d’une succession d’impressions en impression de succession ? Les neurosciences ne voient rien à objecter à ce qu’une séquence d’événements soit représentée par une séquence d’activités neuronales dans la mesure où les termes de ces séquences peuvent être mis en corrélation univoque :« Les ensembles de neurones de l’hippocampe représentent la séquence – c’est-à-dire l’organisation temporelle – des événements dans la mémoire. Cela inclut la mémoire des séquences de lieux traversés, les stimuli séquentiels et les séquences d’actions (H. Eichenbaum, Nature, 15, 2014) ».
La représentation d’une séquence peut parfaitement être une séquence de représentations tant que “représentation” veut dire un état mental ou un pattern d’activité cérébrale. Mais ce qui est peut-être valable pour cette représentation désubjectivée ne l’est pas pour l’acte de conscience qui consiste à se représenter, un acte qui ressaisit en l’unité intentionnelle d’un objet temporel le déploiement séquentiel de ses phases successives. La corrélation aperçue par l’expérimentateur entre la suite des phases de la course du rat sur place et une séquence de champs de décharge neuronale demande à être décodée en interne à une étape de traitement de l’information cognitive ultérieure à celle du système hippocampique, en particulier au niveau du cortex préfrontal où s’élaborent les décisions. L’expérience du cours du temps, acte de conscience intégrant la séquentialité externe ou interne sous-jacente, n’a pas dans l’hippocampe un fondement suffisant sans la suite des étapes de traitement. Ce qui invite à la prudence devant la tendance des spécialistes de l’hippocampe à interpréter sa fonction comme celle de l’organisation temporelle des événements dans la mémoire épisodique, ce qui en fait un organe du temps. Pour être validé comme base neurale de la temporalité subjective l’hippocampe doit être réinséré dans le circuit complet où sa contribution prend un sens de temps. Sans cela, le concept de temps est employé par abus de langage à ce niveau.
Des corrélats pour les horizons de rétention et de protention
Le double horizon de rétention du juste passé et de protention vers le futur présente le caractère d’irréalité de l’intentionnel, tandis que les patterns d’activité des neurones sont des réalités actuelles : une corrélation est-elle possible ? De ce que le fonctionnement cérébral n’est pas enfermé dans la réalité actuelle témoignent les phénomènes de répétition des patterns d’activité des neurones de lieu. Répétition rétrospective (replay) : pendant une pause les cellules de l’hippocampe qui avaient successivement déchargé dans l’ordre des places visitées par le rat déchargent à nouveau dans le même ordre mais à un rythme accéléré (Wilson & McNaughton, Science 265, 1994). Mais aussi répétition prospective (preplay) : pendant la pause de la souris entre une piste familière et une piste nouvelle, la suite temporelle des décharges des cellules de lieu correspond à la suite spatiale des places qui vont être visitées dans le futur et non à la suite spatiale des places qui ont été visitées par le passé (Dragoi & Tonegawa, Nature 469, 2011). Le système replay – preplay à la base de l’extension de l’acte de la conscience-temps au-delà du maintenant actuel vers l’horizon rétentio-protentionnel ? Si la répétition rétrospective de l’activité d’une séquence de cellules de lieu de la course sur la piste familière peut s’expliquer par la réactivation d’une trace cérébrale : réalité actuelle, ce n’est pas le cas pour la répétition prospective. En effet, les cellules qui s’activent en séquences temporelles pendant un repos entre le bout de la piste familière et l’entrée de la piste nouvelle ne sont pas les anciennes cellules de lieu de la piste familière, mais bien les futures cellules de lieu de la piste nouvelle. Devant cette évidence, on ne peut faire autrement qu’en appeler à des mobiles internes de l’organisme, qui contrebalancent l’influence des stimulations externes dans la dynamique de l’auto-organisation séquentielle des assemblées cellulaires de l’hippocampe pour, ou dans l’expectative, ou en prévision de l’expérience future. Le phénoménologue ne manquera pas de souligner ici un geste du naturalisme en direction de l’intentionnalité de la conscience protentionnelle.
Sans vouloir sous-estimer les affinités qu’on a signalées, une sérieuse limitation à la possibilité du dialogue reste que la phénoménologie de la conscience du temps n’est pas un modèle théorique en attente de validation empirique, mais un effort, – mieux : une invitation à faire effort pour remonter de l’évidence de notre expérience des objets temporels aux conditions nécessaires a priori de leur constitution. En retournant le regard de l’attention vers les sources subjectives de l’ordre linéaire et univoque des moments de la durée d’un objet ou d’un événement actuellement perçu, on tente de ressaisir le sens d’être pour un sujet percevant de l’identité permanente de cet objet ou événement à travers la variété de ses phases successives. Appliqué dans la rigueur de la voie husserlienne, ce procédé se singularise par le fait que le phénoménologue prend résolument position dans l’actualité du “maintenant” présent de la conscience perceptive. La structure rétentionnelle de cette conscience rend possible le rappel, – ou plutôt le maintien en prise, directe ou indirecte – des phases écoulées de la durée de l’objet temporel, comme horizon de l’acte de perception actuel. La constante possibilité d’actes de visée attentionnelle pénétrant les horizons de rétention des maintenant successifs de l’apparition de l’objet déploie la forme structurale de linéarité continue unidirectionnelle, condition a priori de sa présentation à la conscience. Cette structure s’étend rétrospectivement sur la durée entière de l’objet jusqu’au moment initial. Mais les flèches de visée intentionnelle qui la traversent émanent d’un foyer unique, qui est le présent vivant de l’acte de perception actuel. En vain cherchera-t-on un pareil privilège d’une phase particulière de « maintenant actuel » dans l’ordre séquentiel des assemblées cellulaires recrutées pour l’encodage du temps de course. Simplement, la dynamique intrinsèque du réseau est telle que les sélectivités d’activation des cellules se distribuent automatiquement de manière à couvrir la totalité sans lacunes des phases temporelles.
La phénoménologie de la conscience du temps veut ressaisir par la réflexion seule les conditions de possibilité subjectives de notre expérience du temps. La recherche empirique apporte avec la découverte des neurones de temps sa contribution à la détermination des bases biologiques de notre expérience du temps. La grande différence des deux approches n’encourage guère le dialogue. Cependant, il est peu satisfaisant que le vécu du passage du temps plane au-dessus d’un néant – ou d’un chaos – cérébral dans l’être de la nature qui éprouve ce vécu. Aussi peu satisfaisant qu’une science des bases cérébrales du temps qui négligerait de prolonger la hiérarchie des étapes de traitement de l’information cognitive jusqu’à rejoindre l’expérience consciente du temps qu’elle prétend expliquer. Lançant une passerelle entre les deux rives de la phénoménologie et des neurosciences cognitives, on s’est demandé dans cet essai si le diagramme de la constitution du temps dans la conscience ne pourrait pas s’interpréter comme l’équivalent d’un système de géométrie interne pour la représentation cartographique des suites d’événements de l’expérience sur des cartes cognitives combinant des dimensions non limitées à l’espace physique et au temps chronométrique. Sans chercher un corrélat cérébral du sujet constituant le temps, l’analogie de la donation de sens au temps écoulé et à l’espace perçu par les actes de conscience du sujet percevant nous oriente vers le probable réemploi d’une même carte cognitive dans l’hippocampe pour le temps écoulé et pour l’espace parcouru. Les mêmes cellules qu’active dans le même ordre la course du rat dans le labyrinthe s’activent également lorsque le rat est contraint de courir sur place : une circonstance dans laquelle les pics d’activité successifs des neurones de lieu semblent ne devoir représenter que les phases successives du temps de course. Auquel cas les neurones de lieu fonctionnent aussi comme neurones de temps. Sans négliger que ce recodage de l’information cognitive du mode spatial au mode temporel ne requiert apparemment ni conscience ni réflexion, on peut au moins noter avec intérêt que les ressources mobilisées par le vivant pour “faire sens” avec la durée sont empruntées à ses moyens de repérage dans l’espace. Comme sur le plan de la conscience réflexive d’après Husserl, une géométrie interne spontanée sous-tend le cours du temps vécu.
La fixation de l’absolue singularité de l’événement en contexte par la mémoire épisodique résulterait d’un processus complexe de spatialisation de l’expérience. De la séparation des patterns d’activité de cellules voisines ou distantes du cortex cérébral émergent les cartes cognitives. La mise en registre de ces cartes – spatiales et non-spatiales – requiert de nombreux changements de référentiels et la solution des problèmes mathématiques de dualité des géométries modèles, euclidiennes ou non euclidiennes (Bennequin et al. 2009). Un processus de neuro-computation qui s’effectue à tous les niveaux d’organisation du centre à la périphérie du cerveau et vice versa, entre les topies (rétinotopie, tonotopie, somatotopie) des aires sensorielles et les répertoires moteurs des actions. Autorisant une description des conditions a priori de notre expérience du temps qui sauvegarde la neutralité formelle du schématisme transcendantal, les diagrammes de la conscience du temps laissent dans l’indétermination la question du substrat cérébral. Que ces conditions puissent un jour être fondées en nature par une bio-géométrie de la dynamique du fonctionnement cérébral est une éventualité à prendre au sérieux pour une naturalisation de la phénoménologie du temps.